MicroRNAs as biomarkers for the diagnosis and prognosis of human cancer
Daniela Asslaber, Josefina Piñón Hofbauer, Richard Greil, Alexander Egle
Laboratory for Immunological and Molecular Cancer Research, IIIrd Medical Department with Hematology, Medical Oncology, Hemostaseology, Rheumatology and Infectiology of the Paracelsus Medical University Salzburg, Müllner Hauptstrasse, Salzburg, Austria
Correspondence: Alexander Egle, Laboratory for Immunological and Molecular Cancer Research, 3rd Medical Department at the Paracelsus Medical University Salzburg, Austria, Müllner Hauptstrasse 48 5020 Salzburg.
E-mail:a.egle@salk.at
Key words: Micro RNA, cancer, biomarker, chronic lymphocytic leukemia.
Received for publication: 23 October 2010.
Revision received: 1 December 2010.
Accepted for publication: 1 December 2010.
©Copyright D. Asslaber et al., 2010
Licensee PAGEPress, Italy
Journal of Nucleic Acids Investigation 2010; 1:e14
doi:10.4081/jnai.2010.e14
Abstract
miRNAs are small-noncoding RNA molecules that regulate gene expression on a posttranscriptional level. A number of oncogenes and tumor suppressors were found to be targets of miRNAs and global miRNA expression signatures were able to distinguish between cancerous and non-cancerous tissues. Therefore it was not surprising that some miRNAs could be linked to the pathogenesis of cancer. In this review we provide an overview of the use of microRNAs as diagnostic and prognostic tools in cancer and focus on the use of miRNA expression as biomarker for disease activity.
|
Introduction
Although miRNAs were already discovered in the early 90s1 they did not become the focus of cancer research until the pioneering work of Calin and colleagues was published in 2002.2 The great novelity of this work was the finding that miR-15 and miR-16 are encoded on chromosome 13q14 in B-cells derived from patients with chronic lymphocytic leukemia (CLL). The 13q14 locus is frequently deleted in human CLL (>50%) and associated with a good prognosis which led the authors to the hypothesis that miRNAs might be involved in the pathogenesis of human cancers. Starting with this report, the number of publications on miRNAs listed in Pubmed has nearly exponentially increased with every year. Nowadays, there is no doubt that miRNAs not only represent novel tools for the characterisation of human cancers that might help to better understand cancer development and maintenance, but also offer new therapeutic options. As a result, one novel treatment concept is the miRNA replacement therapy that introduces miRNA mimics to restore miRNA function in cancers that are dysfunctional for this miRNA.3 This strategy might offer new perspectives in cancer treatment in the future. Furthermore, there is evidence to suggest that microRNAs are not only involved in tumor biology but can also serve as predictors for cancer progression or therapy response, and are biomarkers for the activity of pathways important in tumorigenesis as has been reported by our group. The importance of miRNAs in this respect and the future perspectives in clinical practice are reviewed here.
miRNAs and human cancer
MicroRNAs are small non-protein coding RNA molecules that regulate gene expression on a post-transcriptional level via binding to the 3' UTR of mRNA targets thus resulting in mRNA degradation or repression.4,5,6,7,8 More than 5300 human genes are thought to be regulated by miRNAs, amounting to about 30% of all genes and over 60% of all protein-coding genes being known miRNA targets. Many of these miRNAs are expressed in a highly tissue-specific manner and contribute to the establishment and/or maintenance of the characteristic tissue-related gene expression signature.9,10,11,12 Moreover, miRNAs were shown to regulate the fate of cell lineages as nicely demonstrated for the hematopoietic system. During hematopoiesis every step from the multipotent progenitor to mature myeloid and lymphoid cells is regulated by miRNAs.13
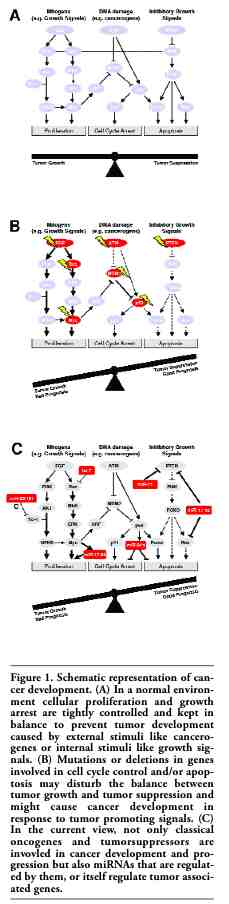
It is not suprising that miRNAs also play a major role in cancer development and progression. The commonly accepted view is that cancer results from a disturbed balance between factors that either promote or suppress tumor formation.14 Important oncogenes that are frequently involved in carcinogenesis such as Ras or c-Myc deliver growth signals and increase cellular proliferation or disable tumor suppressor signals (like MDM2), while tumor suppressors like p53 or PTEN counteract these stimuli. For this review we have used a restricted set of cancer-related genes in order to create a graphical representation of the potential roles of miRNA in the establishment of a cancer phenotype. The choice of displayed genetic targets was influenced by the availability of data on miRNA interaction. Needless to say, such a representation greatly oversimplifies the picture, however this illustration is designed to demonstrate proof of concepts rather than create an adequate picture of real transformation processes (Figure 1A) shows a schematic representation of the chosen oncogenic and tumor suppressor genes and their proposed interaction on the levels of mitogenic response or DNA damage. After achieving mutations in these genes and/or after enhanced production of growth signals or mutations in the corresponding signal transduction molecules the imbalance between cellular growth and growth suppression can be disturbed and tumor cells might be generated (Figure 1B). In already established tumors, alterations in these genes might also contribute to a bad prognosis as shown for p53 and ATM loss of function aberrations or increased MDM2 activity in (see below). A novel concept now integrates miRNAs in this tumor formation concept and shows that an aberrant expression of key miRNAs that regulate those oncogenes and tumorsuppressors might also contribute to tumor formation (Figure 1C).
miRNAs as biomarkers in cancer
Nearly every human cancer has been extensively microarrayed in recent years. Although, the biological readout of such miRNA signatures, that do not often correspond to each other, might be debatable, it was clear already very early that miRNA expression clearly differs between healthy and malignant tissue15 and that several oncogenes and tumor suppressors are targets of miRNAs. Therefore, it was an obvious issue to analyse the diagnostic, prognostic and predictive value of microRNAs in human cancer.16,17 In the following section we will focus on i) the emerging use of circulating miRNAs in cancer diagnosis, ii) two selected miRNAs with an impact on cancer progression, as paradigm for oncogenic miRNAs (miR-21) and tumorsuppressive miRNAs (let-7), and iii) review the role of miR-34a as biomarker for p53 activity, as a prototypic model for a new way to exploit information from miRNA expression studies.
Circulating miRNAs as biomarkers for cancer diagnosis and metastasis
It was demonstrated, recently, that the signatures of circulating miRNAs can be informative in cancer diagnosis. The first evidence that miRNAs expressed in various tissues might enter the blood stream and remain detectable came from Lawrie et al.18 The authors found that the tumor-associated miRNAs miR-155, miR-210 and miR-21 were increased in serum from diffuse large B-cell lymphoma patients as compared to healthy controls.18 Using a human prostate cancer xenograft mouse model Mitchell et al. could validate the technique of cancer classification by miRNA serum level detection.19 Subsequent studies found differential miRNA expression in serum and plasma of lung, ovarian and colorectal cancer patients as compared to healthy individuals.20,21,22 Also for breast cancer a panel of circulating differentially expressed miRNAs was discovered.23,24,25 Among those miR-21 was identified,26 a miRNA that is frequently deregulated in various types of cancer and often related to cancer progression and prognosis as described in the section below. Not only in solid tumors, but also in leukemias, circulating miRNAs might be useful as biomarkers for cancer diagnosis. In the case of AML the miR-92a/miR-638 ratio in blood serum could best discriminate between leukemic and healthy blood.27 Such a technology may be especially helpful for the diagnosis of leukemia in patients displaying a so-called aleukemic presentation of AML, where the tumor cells are not present in the peripheral blood, but only in the bone marrow.
Moreover, circulating miRNAs may be informative regarding metastasis. In prostate cancer several circulating miRNAs were correlated to the occurence of metastatic versus localized tumors, with mir-375 and miR-141 as the most pronounced markers for high-risk tumors in general.28 Also for breast cancer a very recent publication demonstrated the correlation of certain miRNA (miR-34a and miR-155) serum levels to the presence of metastasis.29 If the diagnostic impact of certain circulating miRNAs can be validated in the future and in large patient cohorts, this would offer the great opportunity to detect cancer and/or metastasis very early during blood routine examiniations, without the need for biopsies, and with a greater chance for successful treatment for each individual patient.
miRNAs as Biomarkers for cancer progression
The first report that linked cancer progression and miRNA expression came from Calin et al.30 The authors found a miRNA signature that discriminated between high and low risk CLL patients. Subsequent analysis revealed a number of further miRNAs that were correlated to disease progression in a great variety of different human cancers, which was excellently reviewed by Croce et al.31,32 In this section we will therefore just focus on let-7 as paradigm for a tumorsuppressive miRNA and miR-21 as paradigm for an oncogenic miRNA.
Let-7
Let-7 has become a prototype for a miRNA that functions as a tumor suppressor as it regulates the expression of the Ras oncogene frequently mutated in cancer.33 Downregulation of let-7 may lead to overexpression of Ras and thus constitutive active prosurvival and pro-proliferation signalling cascades that lead to cancer (Figure 1C). Indeed, it was reported that low let-7e34 and let-7-2a35 expression were associated with decreased overall survival in squamous cell carcinoma34 and adenocarcinoma35 patients, respectively, as well as with shortened postoperative survival.36 In human glioblastoma and prostate cancer cells let-7a replacement therapy has led to decreased proliferation and migration in vitro and reduced tumor growth in vivo.37,38 Furthermore, it was established that let-7 was one of 5 miRNAs (miR-25, miR-191, let-7e, miR-34c-5p and miR-34a) that could discriminate between the occurrence of squamous cell carcinoma and adenocarcinoma, two subtypes of non-small cell lung cancer.34 These data suggest that let-7 can serve both as a biomarker for lung cancer subtypes as well as a prognostic marker that predicts disease progression (and response to treatment).
miR-21
The prototype of an oncogenic miRNA that was frequently found to be overexpressed in human cancer39,40,41,42,43,44,45,46 (prostate, stomach, breast, pancreatic, colon, glioblastoma and squamous cell carcinoma) is miR-21. This miRNA was shown to regulate a variety of tumor suppressors such as Pdcd4,47,48 Cdc25A,49 tropomyosin,50 BTG251 and PTEN.52 miR-21 overexpression in vitro has led to increased proliferation, invasion, migration, metastasis and chemoresistance in pancreatic, ovarian, colon and hepatocellular cancer cells.41,47,52,53,54 Knockdown of miR-21 on the other hand could reconstitute normal proliferation and invasion rates47,51,54 and induced apoptosis.45,46 In vivo elevated levels of miR-21 were associated with advanced tumor stages and the occurrence of metastasis in colon44 and breast cancer.55,56 In head and neck,46 as well as lung cancer,40 high miR-21 expression could predict shorter overall survival times. The important role of miR-21 in cancer development and progression was reported by Medina et al.57 who generated a mouse model conditionally expressing miR-21. These mice developed a pre-B malignant lymphoid-like phenotype including lymphadenopathy and splenomegaly. After miR-21 inhibition tumor cells rapidly died upon apoptosis and tumors regressed in between a few days.57 A similar study was provided by Hatley et al.58 The authors generated miR-21 gain- and loss- of function mice and found enhanced tumorigenesis in miR-21 gain-of function mice, while miR-21 loss-of function mice were partially protected against tumor formation.58
In conclusion, miRNAs can act as tumor suppressors as well as in an oncogene-like manner. In these ways they seemingly can influence disease outcome and may be useful markers to predict survival times. Development of targeting strategies for those miRNAs with distinct contributions to pathogenesis will be very interesting.
miRNAs as Biomarkers for gene activity
Not all miRNAs displaying prognostic power in human cancer necessarily have a unequivocal role in disease pathogenesis. It may also be possible that miRNAs may be useable as readouts of the complex genetic pathways involved in cancer and that assessment of their quantity may allow to define interesting surrogate markers for tumor behaviour. We present the example of miRNA-34a in CLL. A clearly defined individual role of this miRNA in the pathophysiology of CLL has not yet been elucidated, but the information yielded by its analysis still yields extremely valuable information. We use the example as a paradigm for additional future exploration in the field of miRNAs.
miR-34a is a reporter of p53 pathway activity
miR-34a belongs to the miR-34 family that consist of two further members: miR-34b and miR-34c. While miR-34a is encoded on chromosome 1 (1p36), mir-34b and c share one transcript on chromosome 11 (11q23), and are expressed in a more tissue specific manner compared to miR-34a. In 2007 five groups in parallel could show that miR-34a is regulated by p53 on a transcriptional level.59,60,61,62,63 miR-34a induction upon DNA damage and oncogenic stress was highly dependent on the presence of functional p53, which binds to evolutionary conserved binding sites upstream of the miR-34a coding sequence.59,60,61,62,63 Recently, a direct connection between miR-34a expression levels and aberrations in p53, namely del17p13 and p53 mutations, could be drawn in CLL.64,65,66,67
It is well established that p53 is one of the most important cancer associated genes and this is reflected in the fact that 50% of all human cancers show deletions or mutations in p53. The tumor suppressor role of p53 is mediated via its central position in signalling pathways that induce apoptosis, DNA damage response, and stop cellular proliferation (Figure 1B). In CLL approximately 5-12 % of patients carry either mutations or deletions in p53.68,69 Both defects are strong independent prognostic markers and associated with a worse overall and treatment free survival.68,70 Moreover, CLL patients with p53 mutations or deletions are often refractory to standard chemotherapy e.g. fludarabine, which is reflected in a relative increase of p53 del/mut patients to 40% in the fludarabine refractory patient group.71 The significantly inferior efficacy of even the currently best chemoimmunotherapy regimes (CLL8), and the fact that some treatment modalities (such as Alemtuzumab) may have a relatively better rate of response even in this high risk group, makes the identification of patients that harbor p53 defects of particular importance in clinical practice.72,73
However, not only direct defects in p53 strongly affect disease progression and therapy response, but also indirect ones, as described for the p53 upstream target ATM, which is induced in response to DNA damage, or the negative p53 inhibitor MDM2, which controls p53 degradation via the proteasome in unstressed cells. The impact of both genes have been described in CLL. Deletions and mutations of ATM (del11q) have a similarly poor prognosis and response to therapy as defects in p53 itself.69,74,75 Similarly, increased MDM2 transcriptional activity due to the SNP-309 polymorphism in the MDM2 promoter76 leading to high p53 degradation in stressed and unstressed cells, indeed negatively influences overall- and treatment-free survival in CLL as has been described by our laboratory.77
This, however, means that 5 independent analyses (p53 deletion, p53 mutation, ATM deletion, ATM mutation, SNP-309) would be needed to predict whether an individual CLL patient belongs to a high risk group with respect to defects along the p53 axis and thus should be treated accordingly (Figure 2).
In our report we established miR-34a to be an effective and reliable surrogate marker for p53 activity.67,78 Interestingly, nearly all CLL patients that were classified as having low miR-34a expression showed defects along the p53 axis, i.e. they were also positive for p53 mutation, p53 deletion, ATM deletion, or the SNP-309 in the MDM2 promoter. The predictive value of miR-34a expression with respect to p53 dysfunction was therefore calculated to approximately 90%. Furthermore, the single analysis of miR-34a expression as a prognostic marker could effectively predict treatment-free survival intervals accurately and with a power that was comparable to the combination of the multiple FISH, PCR, and sequencing assays generally used to establish the status of the p53 axis. In summary our work suggested, that miRNA 34a could be used as a reliable biomarker for p53 pathway integrity in p. Given that clinically information about the integrity of the p53 response in CLL is thought to be the most important factor that determines treatment outcome and survival, a prospective analyses validating of the power of miRNA-34a determination in predicting prognosis and treatment outcome is eagerly awaited.
|
Figure 1. Schematic representation of cancer development. (A) In a normal environment cellular proliferation and growth arrest are tightly controlled and kept in balance to prevent tumor development caused by external stimuli like cancerogenes or internal stimuli like growth signals. (B) Mutations or deletions in genes involved in cell cycle control and/or apoptosis may disturb the balance between tumor growth and tumor suppression and might cause cancer development in response to tumor promoting signals. (C) In the current view, not only classical oncogenes and tumorsuppressors are invovled in cancer development and progression but also miRNAs that are regulated by them, or itself regulate tumor associated genes. |
|
|
Figure 2. miR-34a is a biomarker for p53 activity. In CLL miR-34a is expressed at high levels if the p53 pathway is intact. Aberrations along the p53 axis (e.g. deletion/mutation of ATM or p53, SNP-309 in MDM2) that are associated with a bad prognosis cause decreased miR-34a expression. Thus the miR-34a expression level serves as biomarker for p53 activity and is related to bad prognosis in CLL. |
Conclusions
miRNA profiling is proving invaluable in the study of human cancers. The analysis of miRNA expression patterns may aid in the unraveling of individual lesions central to tumorigenesis, as seems to be the case for mir-15/16, Let-7 or miR-21 to name but a few. However, miRNA expression patterns may also reflect the outcome of complex interactions along a more conventional tumor suppressor/ oncogene pathway, thus allowing the definition of parameters that impact that pathway as well. Our work with miR-34a in CLL provides proof of principle for the latter,67,78 thus opening the field for further discovery, clinical application, and ultimately, improved management of disease.
References
1. Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993;75:843-54.[PubMed]
2. Calin GA, Dumitru CD, Shimizu M, et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA 2002;99:15524-9.[PubMed]
3. Bader AG, Brown D, Winkler M. The promise of microRNA replacement therapy. Cancer Res 2010;70:7027-30.[PubMed]
4. Davis BN, Hata A. Regulation of MicroRNA Biogenesis: A miRiad of mechanisms. Cell Commun Signal 2009;7:18.[PubMed]
5. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004; 116:281-97.[PubMed]
6. Kim VN. MicroRNA biogenesis: coordinated cropping and dicing. Nat Rev Mol Cell Biol 2005;6:376-85.[PubMed]
7. Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet 2010;11:597-610.[PubMed]
8. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009;136: 215-33.[PubMed]
9. Lagos-Quintana M, Rauhut R, Yalcin A, et al. Identification of tissue-specific microRNAs from mouse. Curr Biol 2002; 12:735-9.[PubMed]
10. Lim LP, Lau NC, Garrett-Engele P, et al. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005;433:769-73.[PubMed]
11. Babak T, Zhang W, Morris Q, et al. Probing microRNAs with microarrays: tissue specificity and functional inference. RNA 2004;10:1813-9.[PubMed]
12. Liu CG, Calin GA, Meloon B, et al. An oligonucleotide microchip for genome-wide microRNA profiling in human and mouse tissues. Proc Natl Acad Sci USA 2004;101:9740-4.[PubMed]
13. Bhagavathi S, Czader M. MicroRNAs in benign and malignant hematopoiesis. Arch Pathol Lab Med 2010;134:1276-81.[PubMed]
14. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100:57-70.[PubMed]
15. Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature 2005;435:834-8.[PubMed]
16. Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer 2006;6:857-66.[PubMed]
17. Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer 2006;6:259-69.[PubMed]
18. Lawrie CH, Gal S, Dunlop HM, et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br J Haematol 2008;141:672-5.[PubMed]
19. Mitchell PS, Parkin RK, Kroh EM, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci USA 2008;105:10513-8.[PubMed]
20. Resnick KE, Alder H, Hagan JP, et al. The detection of differentially expressed microRNAs from the serum of ovarian cancer patients using a novel real-time PCR platform. Gynecol Oncol 2009;112:55-9.[PubMed]
21. Taylor DD, Gercel-Taylor C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol Oncol 2008;110:13-21.[PubMed]
22. Chen X, Ba Y, Ma L, et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 2008;18:997-1006.[PubMed]
23. Lodes MJ, Caraballo M, Suciu D, et al. Detection of cancer with serum miRNAs on an oligonucleotide microarray. PLoS One 2009;4:e6229.[PubMed]
24. Wang F, Zheng Z, Guo J, Ding X. Correlation and quantitation of microRNA aberrant expression in tissues and sera from patients with breast tumor. Gynecol Oncol 2010;119:586-93.[PubMed]
25. Zhao H, Shen J, Medico L, et al. A pilot study of circulating miRNAs as potential biomarkers of early stage breast cancer. PLoS One 2010;5:e13735.[PubMed]
26. Asaga S, Kuo C, Nguyen T, et al. Direct serum assay for microRNA-21 concentrations in early and advanced breast cancer. Clin Chem 2010 [Epub ahead of print][PubMed]
27. Tanaka M, Oikawa K, Takanashi M, et al. Down-regulation of miR-92 in human plasma is a novel marker for acute leukemia patients. PLoS One 2009;4:e5532.[PubMed]
28. Brase JC, Johannes M, Schlomm T, et al. Circulating miRNAs are correlated with tumor progression in prostate cancer. Int J Cancer 2010;128:608-16[Abstract]
29. Roth C, Rack B, Muller V, et al. Circulating microRNAs as blood-based markers for patients with primary and metastatic breast cancer. Breast Cancer Res 2010; 12:R90.[PubMed]
30. Calin GA, Ferracin M, Cimmino A, et al. A MicroRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N Engl J Med 2005;353: 1793-801.[PubMed]
31. Croce CM. Causes and consequences of microRNA dysregulation in cancer.Nat Rev Genet 2009;10:704-14.[PubMed]
32. Iorio MV, Croce CM. MicroRNAs in cancer: small molecules with a huge impact. J Clin Oncol 2009;27:5848-56.[PubMed]
33. Johnson SM, Grosshans H, Shingara J, et al. RAS is regulated by the let-7 microRNA family. Cell 2005;120:635-47.[PubMed]
34. Landi MT, Zhao Y, Rotunno M, et al. MicroRNA expression differentiates histology and predicts survival of lung cancer. Clin Cancer Res 2010;16:430-41.[PubMed]
35. Yanaihara N, Caplen N, Bowman E, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006;9:189-98.[PubMed]
36. Del Principe MI, Del Poeta G, Venditti A, et al. Clinical significance of soluble p53 protein in B-cell chronic lymphocytic leukemia. Haematologica 2004;89:1468-75.[PubMed]
37. Lee ST, Chu K, Oh HJ, et al. Let-7 microRNA inhibits the proliferation of human glioblastoma cells. J Neurooncol 2010 Jul 7. [Epub ahead of print][PubMed]
38. Dong Q, Meng P, Wang T, et al. MicroRNA let-7a inhibits proliferation of human prostate cancer cells in vitro and in vivo by targeting E2F2 and CCND2. PLoS One 2010;5:e10147.[PubMed]
39. Volinia S, Calin GA, Liu CG, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci USA 2006;103:2257-61.[PubMed]
40. Markou A, Tsaroucha EG, Kaklamanis L, et al. Prognostic value of mature microRNA-21 and microRNA-205 overexpression in non-small cell lung cancer by quantitative real-time RT-PCR. Clin Chem 2008;54: 1696-704.[PubMed]
41. Moriyama T, Ohuchida K, Mizumoto K, et al. MicroRNA-21 modulates biological functions of pancreatic cancer cells including their proliferation, invasion, and chemoresistance. Mol Cancer Ther 2009 May 12. [Epub ahead of print][PubMed]
42. Iorio MV, Ferracin M, Liu CG, et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res 2005;65:7065-70.[PubMed]
43. Dillhoff M, Liu J, Frankel W, et al. MicroRNA-21 is overexpressed in pancreatic cancer and a potential predictor of survival. J Gastrointest Surg 2008;12:2171-6.[PubMed]
44. Slaby O, Svoboda M, Fabian P, et al. Altered expression of miR-21, miR-31, miR-143 and miR-145 is related to clinicopathologic features of colorectal cancer. Oncology 2007;72:397-402.[PubMed]
45. Chan JA, Krichevsky AM, Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res 2005;65:6029-33.[PubMed]
46. Li J, Huang H, Sun L, et al. MiR-21 indicates poor prognosis in tongue squamous cell carcinomas as an apoptosis inhibitor. Clin Cancer Res 2009;15:3998-4008.[PubMed]
47. Asangani IA, Rasheed SA, Nikolova DA, et al. MicroRNA-21 (miR-21. post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008;27:2128-36.[PubMed]
48. Frankel LB, Christoffersen NR, Jacobsen A, et al. Programmed cell death 4 (PDCD4. is an important functional target of the microRNA miR-21 in breast cancer cells. J Biol Chem 2008;283:1026-33.[PubMed]
49. Wang QZ, Xu W, Habib N, Xu R. Potential uses of microRNA in lung cancer diagnosis, prognosis, and therapy. Curr Cancer Drug Targets 2009;9:572-94.[PubMed]
50. Zhu S, Si ML, Wu H, Mo YY. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1). J Biol Chem 2007;282:14328-36.[PubMed]
51. Liu M, Wu H, Liu T, et al. Regulation of the cell cycle gene, BTG2, by miR-21 in human laryngeal carcinoma. Cell Res 2009;19:828-37.[PubMed]
52. Meng F, Henson R, Wehbe-Janek H, et al. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007;133:647-58.[PubMed]
53. Lou Y, Yang X, Wang F, et al. MicroRNA-21 promotes the cell proliferation, invasion and migration abilities in ovarian epithelial carcinomas through inhibiting the expression of PTEN protein. Int J Mol Med 2010;26:819-27.[PubMed]
54. Zhu S, Wu H, Wu F, et al. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res 2008;18:350-9.[PubMed]
55. Yan LX, Huang XF, Shao Q, et al. MicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008;14:2348-60.[PubMed]
56. Qian B, Katsaros D, Lu L, et al. High miR-21 expression in breast cancer associated with poor disease-free survival in early stage disease and high TGF-beta1. Breast Cancer Res Treat 2009;117:131-40.[PubMed]
57. Medina PP, Nolde M, Slack FJ. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature 2010;467:86-90.[PubMed]
58. Hatley ME, Patrick DM, Garcia MR, et al. Modulation of K-Ras-dependent lung tumorigenesis by MicroRNA-21. Cancer Cell 2010;18:282-93.[PubMed]
59. Chang TC, Wentzel EA, Kent OA, et al. Transactivation of miR-34a by p53 broadly influences gene expression and promotes apoptosis. Mol Cell 2007;26:745-52.[PubMed]
60. Raver-Shapira N, Marciano E, Meiri E, et al. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Mol Cell 2007 Jun 8;26:731-43.[PubMed]
61. Tarasov V, Jung P, Verdoodt B, et al. Differential regulation of microRNAs by p53 revealed by massively parallel sequencing: miR-34a is a p53 target that induces apoptosis and G1-arrest. Cell Cycle 2007;6:1586-93.[PubMed]
62. Tazawa H, Tsuchiya N, Izumiya M, Nakagama H. Tumor-suppressive miR-34a induces senescence-like growth arrest through modulation of the E2F pathway in human colon cancer cells. Proc Natl Acad Sci USA 2007;104:15472-7.[PubMed]
63. He L, He X, Lim LP, et al. A microRNA component of the p53 tumour suppressor network. Nature 2007;447:1130-4.[PubMed]
64. Zenz T, Mohr J, Eldering E, et al. miR-34a as part of the resistance network in chronic lymphocytic leukemia. Blood 2009;113: 3801-8.[PubMed]
65. Mraz M, Malinova K, Kotaskova J, et al. miR-34a, miR-29c and miR-17-5p are downregulated in CLL patients with TP53 abnormalities. Leukemia 2009 23:1159-63[PubMed]
66. Dijkstra MK, van Lom K, Tielemans D, et al. 17p13/TP53 deletion in B-CLL patients is associated with microRNA-34a downregulation. Leukemia 2009;23(3.:625-7.[PubMed]
67. Asslaber D, Pinon JD, Seyfried I, et al. microRNA-34a expression correlates with MDM2 SNP309 polymorphism and treatment-free survival in chronic lymphocytic leukemia. Blood 2010;115:4191-7.[PubMed]
68. Zenz T, Krober A, Scherer K, et al. Monoallelic TP53 inactivation is associated with poor prognosis in chronic lymphocytic leukemia: results from a detailed genetic characterization with long-term follow-up. Blood 2008;112:3322-9.[PubMed]
69. Dohner H, Stilgenbauer S, Benner A, et al. Genomic aberrations and survival in chronic lymphocytic leukemia. N Engl J Med 2000;343:1910-6.[PubMed]
70. Dohner H, Stilgenbauer S, Dohner K, et al. Chromosome aberrations in B-cell chronic lymphocytic leukemia: reassessment based on molecular cytogenetic analysis. J Mol Med 1999;77:266-81.[PubMed]
71. Zenz T, Habe S, Denzel T, et al. Detailed analysis of p53 pathway defects in fludarabine-refractory chronic lymphocytic leukemia (CLL): dissecting the contribution of 17p deletion, TP53 mutation, p53-p21 dysfunction, and miR34a in a prospective clinical trial. Blood 2009;114:2589-97.[PubMed]
72. Keating MJ, Flinn I, Jain V, et al. Therapeutic role of alemtuzumab (Campath-1H) in patients who have failed fludarabine: results of a large international study. Blood 2002;99:3554-61.[PubMed]
73. Lozanski G, Heerema NA, Flinn IW, et al. Alemtuzumab is an effective therapy for chronic lymphocytic leukemia with p53 mutations and deletions. Blood 2004;103: 3278-81.[PubMed]
74. Austen B, Skowronska A, Baker C, et al. Mutation status of the residual ATM allele is an important determinant of the cellular response to chemotherapy and survival in patients with chronic lymphocytic leukemia containing an 11q deletion. J Clin Oncol 2007;25:5448-57.[PubMed]
75. Austen B, Powell JE, Alvi A, et al. Mutations in the ATM gene lead to impaired overall and treatment-free survival that is independent of IGVH mutation status in patients with B-CLL. Blood 2005;106:3175-82.[PubMed]
76. Bond GL, Hu W, Bond EE, et al. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell 2004;119:591-602.[PubMed]
77. Gryshchenko I, Hofbauer S, Stoecher M, et al. MDM2 SNP309 is associated with poor outcome in B-cell chronic lymphocytic leukemia. J Clin Oncol 2008;26:2252-7.[PubMed]
78. Merkel O, Asslaber D, Pinon JD,et al. Interdependent regulation of p53 and miR-34a in chronic lymphocytic leukemia. Cell Cycle 2010;9:2764-8.[PubMed]
[TOP]